Nutrient Fluxes and Interactions in Terrestrial Ecosystems
- rodayasar
- Sep 17, 2024
- 15 min read
Writer: Roda Yasar
Terrestrial ecosystems are complex systems composed of diverse organisms, including plants, animals, and microorganisms, that coexist and interact within shared environments. These ecosystems play critical roles in regulating biogeochemical cycles, influencing processes such as carbon, nitrogen, and water cycles. However, defining the boundaries of these systems can be challenging, as they are often influenced by a variety of factors, including climate, soil composition, and vegetation. This article aims to explore the definition of terrestrial ecosystems, their functions, and their impacts on biogeochemical cycles, thereby providing valuable insights into the complex nature of terrestrial ecosystems.
What is Terrestrial Ecosystems
A critical conceptual consideration in evaluating biogeochemical cycles for terrestrial ecosystems is scale. What are the boundaries of a forest or grassland? Quantitative determinations of fluxes, mass balances, or interactions cannot be made without establishing boundaries, but such pragmatic edges of a terrestrial ecosystem are usually established for the convenience of the investigation rather than to delimit integral functional units. Lakes, streams, or islands appear to have obvious boundaries—but do they? What are the boundaries? How are they affected during periods of flooding or drought? Aquatic invertebrates may be found many meters below the sediment-water interface or horizontally beyond the banks of the stream and “beneath” the terrestrial ecosystem (Hynes, 1974). Where then are the boundaries? Terrestrial landscapes often have political, climatic, topographic, vegetational, or other boundaries. Are these in any way functional boundaries?
The size of “ecosystems” studied in the past has ranged from plots only a few meters in area to the entire planetary surface. The experimental scale has a profound effect on the theoretical context of a functional ecosystem. Larger systems, for example, a global one, tend to be “closed” for mass. As such, they may be in a time-invariant state (equilibrium) or a time-dependent state (disequilibrium). In contrast, smaller ecosystems tend to be “open” concerning mass and may be in a steady-state mode (time-invariant) or a non-steady-state mode (time-dependent). Our comprehension of the processes operating within an ecosystem (Johnson, 1971) is thus framed by the state of the boundaries for the system (open versus closed) and the reaction pathways.
Due to their immense theoretical value, thermodynamic parameters (equilibrium constants, solubility products, phase diagrams) are occasionally used to describe certain natural processes. Implicit in such attempts, however, is that the field conditions must conform to the exacting conditions required by the thermodynamic considerations. More often than not, however, the necessary conditions for thermodynamic equilibrium are not met in the field, and thermal kinetic theory, or some approximation thereof, is required. The kinetic theory represents a reliable body of knowledge equally as powerful as thermodynamics and perhaps more applicable to natural processes.
It is much easier to make quantitative measurements of the process rates in smaller ecosystems, but do such areas exhibit all of the functional properties of a larger system? What are the critical functional properties? What are the linkages with other systems that affect these properties? Such questions are rarely addressed by ecologists or geochemists.
The error limits associated with measurements of rate processes within very large ecosystems may be so great as to compromise any real understanding of the biogeochemistry of these systems. The problem of signal-to-noise ratios is rarely addressed when reporting estimates of rates for processes in ecosystems.
In our biogeochemical studies of the Hubbard Brook Experimental Forest in the White Mountains of New Hampshire (Bormann and Likens 1979), we sought a solution to some of these problems by utilizing rather small, discrete watershed (catchment) ecosystems. We looked for units of the landscape that could be isolated based on flowing water in the system. Because water is such an important vector for chemical flux and cycling in natural forested ecosystems, watershed (phreatic) divides would appear to form both functional and reasonable boundaries, amenable to quantitative analysis (Bormann and Likens, 1967; Likens and Bormann, 1972; Likens et al., 1977; Bormann and Likens, 1979). Our model (Figure 1) accounts for the quantitative movement of matter across the boundaries of such a watershed ecosystem by the three principal vectors-meteorologic (movement by atmospheric forces), geologic (movement by alluvial or colluvial forces), and biologic (movement by animals). We consider the release and storage of materials inside the boundaries, for instance by weathering, or biomass accumulation and release, as internal features of the ecosystem rather than input/output.

Figure 1 A model depicting nutrient relationships in a terrestrial ecosystem. Inputs and outputs to the ecosystem are moved by meteorologic, geologic, and biologic vectors (Bormann and Likens, 1967; Likens and Bormann, 1972). Major sites of accumulation and major exchange pathways within the ecosystem are shown. Nutrients that, because they have no prominent gaseous phase, continually cycle within the boundaries of the ecosystem between the available nutrient, organic matter, and primary and secondary mineral components tend to form an intrasystem cycle. Fluxes across the ecosystem's boundaries link individual ecosystems with the remainder of the biosphere (Likens et al., 1977)
CYCLE INTERACTIONS
Understanding the points at which the major biogeochemical cycles intersect and how these connections are influenced by disturbances is essential. For instance, a fundamental inquiry revolves around the impact of changes in the sulfur cycle on the nitrogen cycle's sensitivity, as well as identifying the most sensitive or pivotal connections between these two cycles. The impact of sulphuric acid in rainfall on nitrification in the soil, the base exchange capacity, and the mineralization activities of microorganisms raises important questions. Although there is limited knowledge about the answers, it is probable that as human activity continues to place strain on the natural environment, the effects on individual biogeochemical cycles will have far-reaching consequences, potentially influencing all interconnected cycles. The composite result may be more important to the overall function of a natural system than an analysis of any single cycle would indicate. It is not clear to us how to obtain such complex answers on a global scale. On the scale of watersheds, however, it is possible by judicious comparative studies or experimental manipulations to elucidate mechanisms, reaction rates, and interactions. Insights may be obtained not only from the results of experimentally imposed disturbance, but it is also possible to “relax” or remove a pre-existing stress on an ecosystem; e.g. reduce the pollution in a stream or stop the cultivation of a field, and observe the recovery of the system. On a global scale, various stress-inducing phenomena are currently occurring, such as heightened CO2 levels in the atmosphere, acid rain resulting from the burning of fossil fuels, and increased use of nitrogenous fertilizers. However, these changes were not deliberately orchestrated as experiments, making it difficult or impossible to create controlled replicates. Given these conditions, it is difficult to measure the impact on each cycle, not to mention the intricate interplay between the cycles. Hence, scientists frequently have resorted to smaller, more manageable systems and simulation models with the hope of extrapolating these results to a global scale. At this point, in the development of such information, however, global valuations of individual biogeochemical cycles are at the very best only rough approximations.
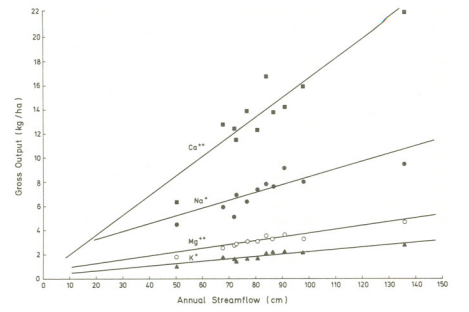
Figure 2 Relationship between streamflow and gross output of cations from 1963 to 1974 at the Hubbard Brook Experimental Forest (Likens et al., 1977)
Hydrologic Cycle
In terrestrial ecosystems, water serves as a crucial connector for chemical elements, playing a pivotal role in various processes and interactions. The hydrologic cycle affects other biogeochemical cycles in numerous direct and indirect ways. Water acts as a solvent, carrier, catalyst, and reagent in many ecosystem reactions and processes. The flux of nutrients into and through many terrestrial ecosystems is largely dependent on the amount and temporal distribution of rain and snow and the chemical content of this water. As an illustration, in an extreme case, (Art et al., 1974) found for a coastal terrestrial ecosystem on Long Island, New York, that all of the inputs of calcium, magnesium, sodium, and potassium came from bulk precipitation and deposition of sea spray. Smaller but significant amounts of these nutrients are added to natural ecosystems in bulk precipitation at greater distances from the ocean (Likens et al., 1977). Likewise, losses of major nutrients are usually a function of the amount of water draining from an area (Figure 2). In this regard, water lost by transpiration, which is an indirect function of the carbon cycle via photosynthesis, is not available for drainage and thereby transpiration serves to reduce the dissolved nutrient flux out of the system. For example, at the Hubbard Brook Experimental Forest, on average, some 38 percent of the annual precipitation is lost as evapotranspiration each year (Figure 3).
The amount of available water affects the rate of primary production (carbon cycle) in terrestrial ecosystems (Leith, 1973) and, hence, the rate of uptake and storage of nutrients in biomass. It also plays a major role in the regulation of the processes of throughfall, leaching, decomposition, and mineralization which are integral parts of all nutrient cycles in humid regions. Water is the primary agent of erosion and transport in humid regions (Bormann et al., 1974; Dunne and Leopold, 1978; Leopoldetalet al., 1964). The Downstream transport of particulate matter constitutes an important loss in terrestrial ecosystems, particularly for relatively insoluble elements such as phosphorus, iron, and lead. A major biogeochemical aspect of erosion is related to the removal of exchange surfaces, since not only does this result in the loss of available nutrients, but the loss of exchange surfaces can greatly alter the capacity of the system to regulate nutrient cycles and to recover from disturbance. The regulation of the hydrologic cycle and, therefore, erosion is dependent on the developmental phase of the terrestrial ecosystem (Bormannand Likens, 1979), and may drastically change by disturbance, man-made or otherwise. It is noteworthy that where knowledge of hydrologic fluxes is uncertain, associated chemical element budgets are also poorly known. Semi-arid and arid ecosystems exemplify this condition. It is difficult at best to deal quantitatively with the biogeochemistry of ecosystems where the hydrologic budget is unknown, unreliable, or ephemeral.
In summary, it is challenging to exaggerate the importance of the water cycle concerning practically every aspect of biogeochemistry. Massive Irrigation works, diversions of major river systems, large-scale cloud seeding, major impoundments, and deforestation or urbanization of catchment areas are all examples of man's conscious or unconscious efforts in manipulating the hydrologic cycle. All of the above have well-documented effects in dislocating or disturbing the biogeochemical activity within the affected area. Perhaps the most extreme hydrological shock happens when the hydrological cycle transitions from its regular state to an ice age state. Could human activities impacting the balance of CO2, atmospheric dust, photochemical smog, and the Earth's albedo lead to such a significant and impactful change?

Figure 3 Relationship among precipitation, streamflow, and evapotranspiration for the Hubbard Brook Experimental Forest from 1965 to 1974. Transpiration accounts for the majority of the annual evapotranspiration water loss for this humid forest ecosystem (Likens et al., 1977).
Carbon Cycle
The carbon cycle often regulates interactions between various other biogeochemical cycles because of its pivotal role in photosynthesis and decomposition. When ecosystem production surpasses decomposition, nutrients are stored as biomass accumulates. However, if decomposition is the dominant process, nutrients are released from the decaying organic matter. It must be realized, however, that these are generalized responses, since actual uptake and mineralization rates will be dependent on the behavior of different vegetational and chemical species, or both (Gosz et al., 1973; Gosz, 1981).
Nutrient release through the decomposition of leaf tissue in the Hubbard Brook Experimental Forest is influenced by critical carbon-to-element or element-to-phosphorus ratios (Gosz et al.,1973). Once the critical C: N, C:P, etc., ratio is attained, mineralization occurs. Threshold ratios may exist for other elements and thereby control their mineralization rates in decomposing organic matter. Thus The presence of one nutrient can affect the availability of another through this type of biogeochemical interaction.
Currently, a great amount of attention is being focused on the carbon cycle because of increasing concentrations of CO2 in the atmosphere from the combustion of fossil fuels and the resulting potential for climatic change (Anonymous, 1977). Spatially, terrestrial ecosystems are quite heterogeneous, some areas store carbon while others lose it. Globally, however, it is thought that terrestrial ecosystems are a net source of carbon for the atmosphere (Olson et al., 1978; Bolin,1977; Woodwell et al., 1978). Although the amounts and assumptions underlying this flux are disputed (Broecker et al., 1979). Moreover, the prevailing wisdom, although not certain either, is that. elevated levels of CO2 in the atmosphere have not, in turn, stimulated terrestrial photosynthesis on a global scale (Woodwell et al., 1978). If net global ecosystem production were enhanced, then storage of nitrogen, phosphorus, and other nutrients in the accumulating biomass would be increased accordingly and transpiration also might be altered.
The so-called “Redfield ratio” for the content of carbon, nitrogen, and phosphorus in organic material is widely quoted and used in mega calculations for ecosystem and global budgets. This atomic ratio for C: N:P or 106:16:1 (weight ratio of 40:7:1) was based on analysis of marine plankton (Redfield, 1958; Redfield et al., 1963). Ratios for terrestrial ecosystems, dead organic matter, or global systems may differ widely from the values for marine plankton (Table 1), and it is not appropriate to use the Redfield ratio generally for terrestrial ecosystems. The fact that the ratios are different is not at all surprising since terrestrial plants have a much greater proportion of carbon (cellulose) stored in structural tissues (e.g. wood). Even 'ocean plants' (Table 1) have a different ratio than that proposed by Redfield because macrophytes and attached algae are included.
(Redfield, 1958) argued convincingly that organisms controlled the proportion of inorganic nitrogen and phosphorus in the sea. Furthermore, he proposed a strong interaction between these two elements which resulted in a net equilibrium and fixed C: N:P ratio in marine organisms. The same argument can be made for terrestrial systems even though the C: N:P ratios are significantly different from those found in the sea. According to Redfield's arguments, phosphorus is the master element that controls the availability of the others, because it is present in the lowest supply at the earth's surface relative to carbon, nitrogen, and oxygen. This relatively simple relationship, however, does not necessarily hold for terrestrial ecosystems where nitrogen, water, and other factors frequently limit growth ( Waringand & Franklin, 1979).
Simulation models of forest growth may give insight into the effects of vegetation disturbance or the effects of accelerated plant growth on biogeochemical cycles (Botkin et al., 1973; Shugart and West, 1977). For example, simulation suggests that catastrophic losses of major tree species(e.g. blights of chestnut, hemlock from the hardwood forests of the eastern USA) would affect biogeochemical cycles in a variety of ways, e.g. via uptake, release, and differential storage of nutrients, gaseous exchange, impaction of aerosols, etc. The carbon cycle would be temporarily affected by such changes, as would be the flux and cycling on, P, S, and other nutrients
Harvesting of forest products by clearcutting provides an example of how various cycles may be affected through their linkages. Immediately following the clearcutting of northern hardwoods in the northeastern USA, photosynthesis would be reduced and decomposition would be increased. As a result, transpiration would be increased; inventories, accumulation, and release of all elements in biomass would be altered depending on the type of harvest and developmental stage of the ecosystem.
Nitrification would be accelerated and there would be increased loss of calcium, magnesium, potassium, sodium, nitrate, phosphate, and hydrogen ions and decreased loss of sulfate in stream water (Bormann and likens, 1979).
Another aspect that should be considered here is the developmental stage of the forest ecosystem (Bormann and Likens, 1979). Carbon flux might be greater for young forests in the early stages of development, but the impaction of sulfurous, nitrogenous, and phosphorus aerosols probably would be less because the architecture (impaction surface) of the forest would not be fully developed. The effects of ecosystem development and dramatic or subtle shifts in species composition on biogeochemical cycles and their interaction are areas worthy of detailed study ( Bormann and Likens, 1979; Gorham et al., 1979).

Table 1 1. Modified from Delwiche and Likens (1977); the amount of phosphorus in land plants decreased 4- fold and ratios were adjusted accordingly; 2. weighted for species composition; 3. weighted for above and below ground materials of different species
Nitrogen Cycle
Annually some 40 Tg of N are fixed commercially as agricultural fertilizer (Delwiche and Likens, 1977). There has been appreciable concern about the role of this fertilizer in the eutrophication of aquatic ecosystems (Kohl et al., 1971), and whether gaseous losses from the soil would be enhanced and thereby produce atmospheric imbalances (McElroy et al., 1977; Crutzen and Ehhalt, 1977; Council for Agricultural Science and Technology, 1976; Hutchinson and Mosier, 1979). E. Lemon and co-workers(personal communication) at Cornell University have found that the vegetational uptake of gaseous NH3 may serve as a balancing mechanism for levels of ammonia in the atmosphere. Agricultural vegetation can release NH3 when ambient atmospheric concentrations are low, and absorb it when gaseous concentrations are high. The uptake rate is accelerated at higher concentrations. Lemon has suggested that if nitrogen were limited to photosynthesis, then losses of NH3-N (from fertilizer as well as naturally-fixed N) in one area could stimulate net production by plants in another area via the mechanism described above and thus influence the storage of atmospheric CO2 in living and dead biomass. In turn, all other major nutrients would be affected proportionally by the changing levels of photosynthetic and decomposition activity.
Biologically significant amounts of nitrogen are added to terrestrial ecosystems each year in rain and snow. At various locations in the eastern US annual inputs in precipitation range between 6 and 10 kg-N ha-l (Likens et al., 1977; Likens, 1972). Wet deposition values in industrialized Europe may be even greater (Soderlund, 1977). Such inputs have increased significantly during the last decade in both the eastern USA (Figure 4) and western Europe (Likens, 1976). There is no quantitative evidence as yet whether these increased inputs have had a significant effect on plant productivity or decomposition in natural ecosystems

Figure 4 Annual nitrate concentrations in precipitation in the northeastern USA (Likens and Bormann, 1980)
Conclusion
Terrestrial ecosystems are complex systems made up of plants, animals, and microbes interacting within their environment. They play a vital role in managing important natural cycles like carbon, nitrogen, and water. However, determining their exact boundaries can be challenging due to varying factors such as climate and soil type.
The size of an ecosystem affects how we study its processes. Large ecosystems are often treated as closed systems, while smaller ones are considered open. This difference can make measuring and understanding ecological processes tricky.
Water is crucial in these ecosystems, affecting nutrient movement and various processes. For example, water loss through evaporation can impact the flow of nutrients.
The carbon cycle is important because it connects with other cycles through photosynthesis and decomposition. Higher CO2 levels can drive climate change and influence these cycles.
The nitrogen cycle is influenced by factors like fertilizer use and its effects on ecosystems and atmospheric balance.
In summary, understanding these cycles and their interactions is key for managing and preserving the environment.
References
Anonymous (1977). Energy and Climate. Washington D.C., National Academy of Sciences.
Art, H. W., Bormann, F. H., Voigt, G. K., and Woodwell, G. M. (1974). Barrier Island Forest Ecosystem: role of meteorologic nutrient inputs. Science. pp. 60-62.
Bolin, B. (1977). Changes of land biota and their importance for the carbon cycle. Science. pp. 613-615.
Bormann, F. H. & Likens, G. E. (1967). Nutrient cycling. Science. pp. 424-429.
Bormann, F. H. & Likens, G. E. (1979). Pattern and Process in a Forested Ecosystem. New York, Springer-Verlag.
Bormann, F. H., Likens, G. E., Siccama, T. G., Pierce, R. S., and Eaton, J. S. (1974). The export of nutrients and recovery of stable conditions following deforestation at Hubbard Brook. Ecol. Monogr. pp. 255-277.
Botkin, D. B., Janak, J. F., and Wallis, J. R. (1973). Estimating the effects of carbon fertilization on forest composition by ecosystem simulation. In Woodwell, G. M. and Pecan, E. V. (eds), Carbon and the Biosphere. Proc. 24th Brookhaven Symposium in Biology. pp. 328-344.
Crutzen, P. J. and Ehhalt, D. H. (1977). Effects of nitrogen fertilizers and combustion on the stratospheric ozone layer. Ambio. pp. 112-117.
Council for Agricultural Science and Technology (1976). Effect of increased nitrogen fixation on stratospheric ozone. Report No. 53. Iowa State University, Ames.
Delwiche, C. C. and Likens, G. E. (1977). Biological response to fossil fuel combustion products. In Stumm, W. (ed), Global Chemical Cycles and Their Alterations by Man. Berlin, Dahlem Konferenzen. pp. 33-88.
Dunne, T. and Leopold, L. B. (1978). Water in Environmental Planning. San Francisco, W. H. Freeman.
Gorham, E., Vitousek, P. M. and Reiners, W. A. (1979). The regulation of chemical budgets throughout terrestrial ecosystem succession. Ann. Rev. Ecol. Syst. pp. 53-84.
Gosz, J. R. (1981). Nitrogen cycling in coniferous ecosystems. In Terrestrial Nitrogen Cycles - Processes, Ecosystem Strategies, and Management Impacts. SCOPE Report 18, Workshop at Gysinge Wärdshus, Österfärnebo, Sweden.
Gosz, J. R., Likens, G. E. and Bormann, F. H. (1973). Nutrient release from decomposing leaf and branch litter in the Hubbard Brook Forest, New Hampshire. Ecol. Monogr. pp. 173-191.
Hutchinson, G. L. and Mosier, A. R. (1979). Nitrous oxide emissions from an irrigated cornfield. Science. pp. 1125-1127.
Hynes, H. B. N. (1974). Further studies on the distribution of stream animals within the substratum. Limnol. Oceanogr. pp. 92-99.
Johnson, N. M. (1971). Mineral equilibria in ecosystem geochemistry. Ecology. pp. 529-531.
Kohl, D. H., Shearer, G. B. and Commoner, B. (1971). Fertilizer nitrogen: contribution to nitrate in surface water in a corn belt watershed. Science. pp. 1331-1334.
Leopold, L. B., Wolman, M. G. and Miller, J. P. (1964). Fluvial Processes in Geomorphology. San Francisco, W. H. Freeman.
Lieth, H. (1973). Primary production: terrestrial ecosystems. Human Ecology. pp. 303-332.
Likens, G. E. (1976). Acid precipitation. Chemical and Engineering News. pp. 29-44.
Likens, G. E. and Bormann, F. H. (1980). The role of watershed and airshed in lake metabolism. In Rodhe, W., Likens, G. E. and Serruya, C. (eds), Proc. Symposium on Lake Metabolism and Lake Management. Arch. Hydrobiol. Beih. Ergebn. Limnol. pp. 195-211.
Likens, G. E., Bormann, F. H., & Johnson, N. M. (1981). Interactions between major biogeochemical cycles in terrestrial ecosystems. In Some perspectives of the major biogeochemical cycles. pp. 93-112.
Likens, G. E., Bormann, F. H., Pierce, R. S., Eaton, J. S. & Johnson, N. M. (1977). Biogeochemistry of a Forested Ecosystem. New York, Springer-Verlag.
McElroy, M. B., Wofsy, S. C. and Yung, Y. L. (1977). The nitrogen cycle: perturbations due to man and their impact on atmospheric N₂O and O₃. Phil. Trans. Roy. Soc. (London) B. pp. 159-181.
O'Sullivan, P. E. (1979). The ecosystem-watershed concept in the environmental sciences - a review. Internat. J. Environ. Studies. pp. 273-281.
Olson, J. S., Pfuderer, H. A. and Chan, Y. -H. (1978). Changes in the global carbon cycle and the biosphere. Oak Ridge National Laboratory, Environmental Sciences Division Publication.
Redfield, A. C. (1958). The biological control of chemical factors in the environment. Amer. Sci. pp. 205-221.
Shugart, H. H. and West, D. C. (1977). Development of an Appalachian deciduous forest succession model and its application to assessment of the impact of the chestnut blight. J. Environ. Management. pp. 161-179.
Söderlund, R. (1977). NO pollutants and ammonia emissions - a mass balance for the atmosphere over Northwest Europe. AmMo. pp. 118-122.
Waring, R. H. and Franklin, J. F. (1979). Evergreen coniferous forests of the Pacific Northwest. Science. pp. 1380-1386.
Woodwell, G. M., Whittaker, R. H., Reiners, W. A., Likens, G. E., Delwiche, C. C. and Botkin, D. B. (1978). The biota and the world carbon budget. Science. pp. 141-146.



Comments